ドキシサイクリンはミトコンドリアのリボソームを阻害してがん細胞の酸化ストレスを高める
ミトコンドリアは太古の昔に細菌が原始真核細胞に寄生してできました。抗生物質のドキシサイクリンとアジスロマイシンは、細菌のリボソームの働きを阻害して抗菌作用を発揮します。細菌のリボソームとミトコンドリアのリボソームは相同性が高いため、これらの抗生物質はミトコンドリアの働きを阻害し、ATP産生を低下させ、活性酸素種の産生量を高めます。ミトコンドリア・リボソームの阻害がフェロトーシス誘導を促進する理由を解説します。
◉ ミトコンドリアは元は細菌だった
ミトコンドリアは赤血球以外の全ての細胞に存在する細胞小器官です。
1個の細胞当たり平均で300~400個のミトコンドリアが存在します。肝臓や腎臓や筋肉や脳など代謝が活発な細胞には数千個のミトコンドリアが存在し、細胞質の40%程度を占めています。体内のミトコンドリアは全部で体重の約10%を占めると言われています。真核細胞のミトコンドリアは好気性細菌のαプロテオバクテリアが原始真核細胞に寄生してできました。まだ酸素が無い太古の地球に生きていた生物は解糖系のみでエネルギーを得ていました。ところが、海中に発生した藻類が光合成によって吐き出す酸素が大気中に増えていくと、酸素の無い環境で生きていた生物は酸化力の強い酸素に触れることでダメージを受けるようになります。そのためこの時期には原始真核生物の多くが絶滅し、あるいは酸素の影響を受けることのない深海などに移動していきました。
このような状況で誕生したのが、酸素を使ってATPを生成する好気性細菌です。そして、約20億年前に好気性細菌のα-プロテオバクテリアが原始真核細胞に寄生して、ミトコンドリアになったと考えられています(下図)。
図:嫌気性(酸素を使わない)の原始真核細胞に好気性細菌のαプロテオバクテリアが食作用で取り込まれて共生するようになりミトコンドリアになった。αプロテオバクテリアに存在していた遺伝子の多くは真核細胞の核内に移動し、ミトコンドリアのタンパク質の多くは核の遺伝情報によって作られるようになった。
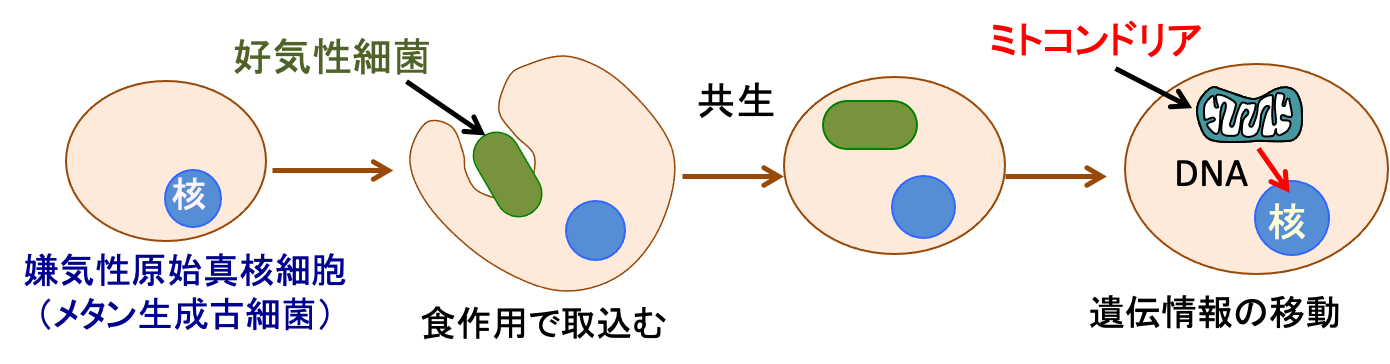
好気性細菌は生体にダメージを与える酸素をグルコース(ブドウ糖)に結合させ、二酸化炭素と水に分解し、さらにその過程でATPを大量に生成することができます。この細胞内共生によって酸素が豊富な環境で生物が急速に進化することになります。
つまり、原始真核生物は、酸素を利用できるα-プロテオバクテリアを細胞内に取り込んで共生することによって、酸素分圧の高い環境で生きていけるようになったのです。
このように、ミトコンドリアはかつて細菌であったため、見かけは細菌に似ています。直径は1ミクロン(1ミクロンは1000分の1ミリ)以下で、長さは1~4ミクロン程度で、俵型やいも虫様の立体構造をしています。
ミトコンドリアは2枚の膜(内膜と外膜)によって細胞質から隔てられ、内膜は複雑に入り組んで「クリステ」という無数の襞や管を形成しています。内膜が襞上にくびれているのは、表面積を増やすためで、この内膜でATPの産生が行われています。
内膜上には、電子伝達系やATP合成にかかわる酵素群などが一定の配置で並んでいます。マトリックスには、TCA回路に関わる酵素やミトコンドリア独自のDNAなどが含まれています。
タンパク質合成のためにリボソームも持っていてミトコンドリア内でタンパク質合成もできます。そして、ミトコンドリア自身が増殖もします。
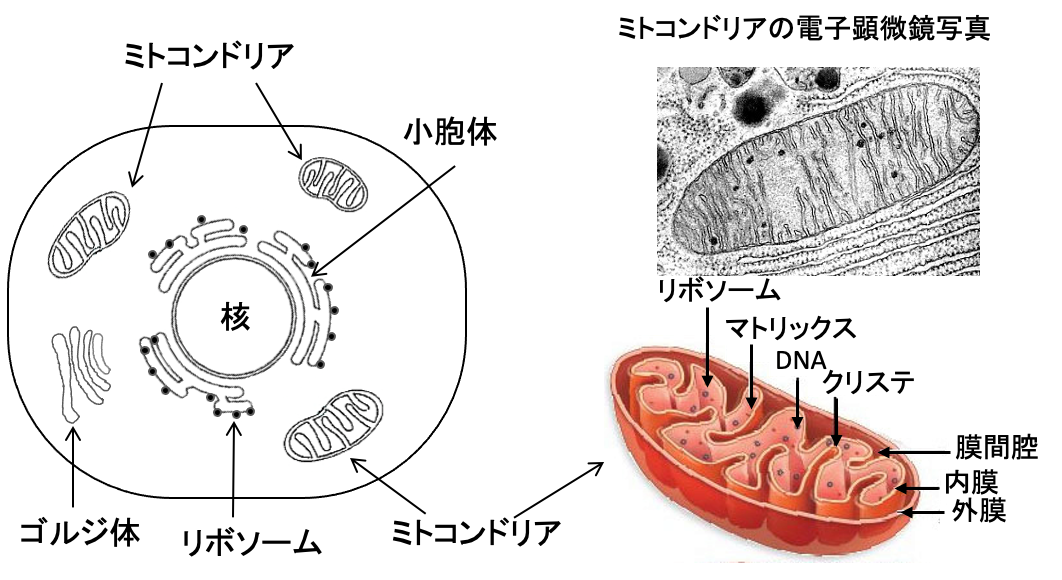
図:細胞内には機能を分担するために様々な小器官が存在する。ミトコンドリアは酸素を使ってグルコースや脂肪酸やアミノ酸を燃焼してATPを産生する働きや、物質代謝やアポトーシスの制御など多彩な機能を担っている。マトリックスにはTCA回路に関わる酵素やミトコンドリア独自のDNAやタンパク質合成のためにリボソームが存在する。
◉ タンパク質はリボソームで作られる
タンパク質を作る遺伝子情報はDNAに並ぶ塩基配列によって保存されています。細胞核内のDNAが転写されてメッセンジャーRNA(mRNA)となり、リボソームで20種類のアミノ酸からなるタンパク質へと翻訳されます。
タンパク質はアミノ酸が複数結合した直鎖状の分子です。リボソームで作られたタンパク質は、小胞体で特定の立体構造に折り畳まれて機能を発揮するようになります。
リボソームはタンパク質を作る細胞構造です。リボソームは細胞質内に浮遊しているか、小胞体に付着しています。細胞内に自由に浮遊しているリボソームは細胞内で利用されるタンパク質を合成しています。小胞体に付着しているリボソームは、細胞内または細胞外で使用されるタンパク質の合成を行います。リボソームが付着した小胞体を粗面小胞体といいます。
リボソームで作られた膜タンパク質や分泌タンパク質は、小胞体内やゴルジ体で「タンパク質の折り畳み」や、「糖鎖の結合」などタンパク質の翻訳後修飾を受けて正しい機能を発揮できるタンパク質として完成します。リボソームは、メッセンジャーRNA(mRNA)、トランスファーRNA(tRNA)、および複数の翻訳因子と協調してタンパク質合成のプロセスを調整する非常に複雑な高分子構造です。大きなサブユニットと小さいサブユニットの2つのサブユニットが結合した構造です。大サブユニットはアミノ酸間にペプチド結合を形成するペプチジルトランスフェラーゼ反応の触媒作用に関与し、小サブユニットはmRNAの結合と解読のプラットフォームを提供します(下図)
図:リボソームは大きなサブユニットと小さいサブユニットが結合した構造をしている。リボソーム内では、mRNAに転写された遺伝情報に従いトランスファーRNA(tRNA)が対応するアミノ酸を連結することでタンパク質を合成する。mRNAのコドン(3文字の塩基配列)が一つのアミノ酸の種類を決定する。翻訳後のタンパク質は3次元的に折り畳まれたり、糖鎖結合などの翻訳後修飾を受けて、機能をもったタンパク質が完成する。
タンパク質の大きさは分子量で比較します。一方、リボソームのような複雑で大きい細胞内分子の大きさの比較は「沈降係数」が用いられます。沈降係数(S値)とは単位加速度あたりの沈降速度で、超遠心機を開発したテオドール・スベドベリ(Theodor Svedberg)にちなむ単位です。
より大きな粒子はより速く沈降し、より大きな沈降係数を有します。 その構造物が複数のサブユニットから構成される場合、沈降係数は相加的ではありません。粒子の質量あるいは体積にのみ依存するわけではなく、2つの粒子が互いに結合している時は表面積の損失が必然的に起こるために、結合粒子の沈降係数は個々の粒子の個別に測定されたスベドベリ値の和にはなりません。リボソームはほとんどの場合、それらの沈降係数によって同定されます。例えば、細菌由来の70Sリボソームは実際に70スベドベリの沈降係数を有しています。一方、70Sリボソームは50Sサブユニットと30Sとサブユニットから構成されており、サブユニットのスベドベリ値の合算よりも小さくなります。
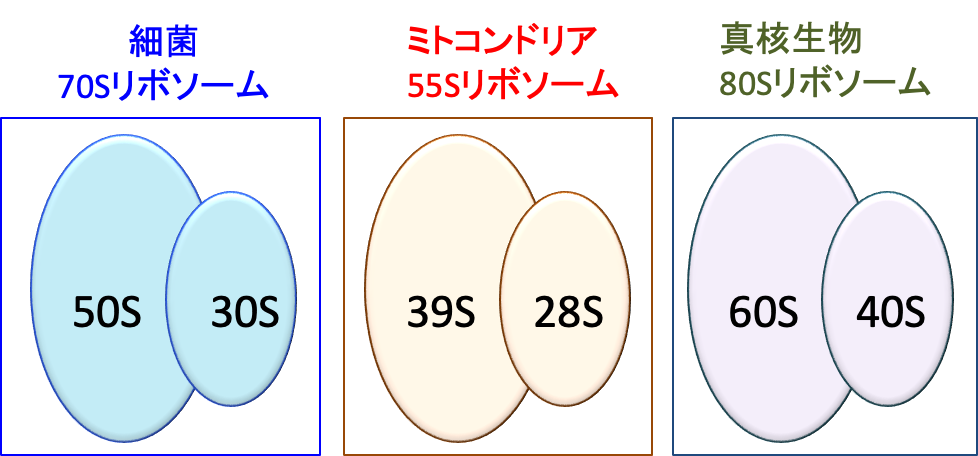
私たち人類やその他の動物、植物、菌類などの真核細胞にあるリボソームは、細菌や古細菌のものより大きく80Sリボソームと呼ばれています。こちらは、小さい方の40Sサブユニットと大きい方の60Sサブユニットで構成されています。
私たちの細胞にあるミトコンドリアは細胞本体が持つものより小さい55Sリボソームを持っていて、細胞質にあるものとは別々に作られます。
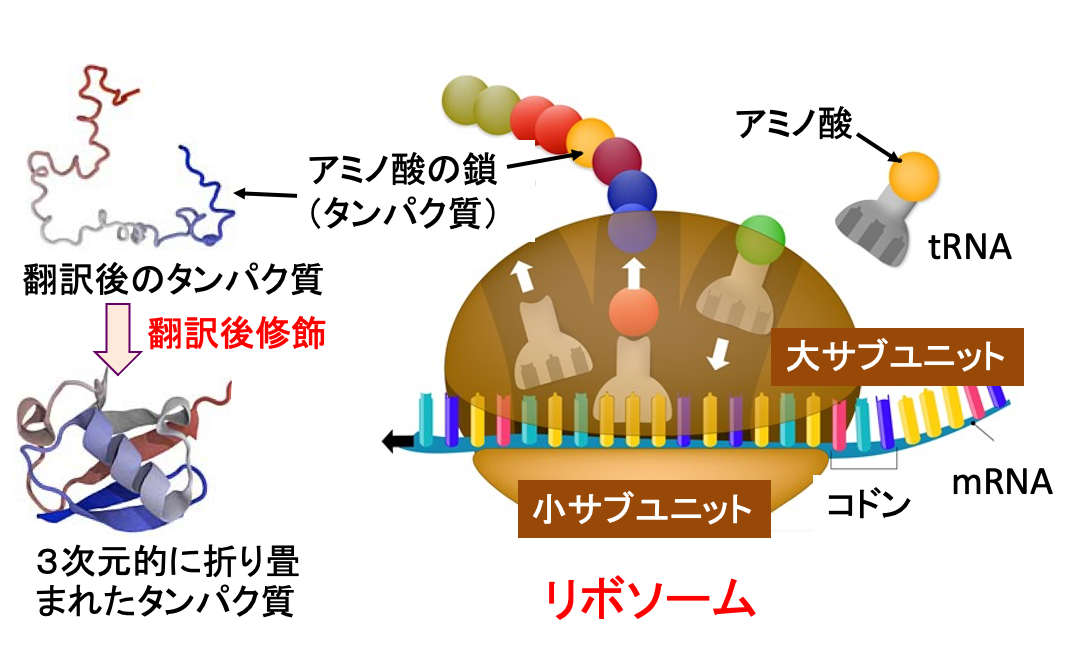
図:リボソームは大小2種類のサブユニットから構成され、真核生物は40Sサブユニットと60Sサブユニットから構成される80Sリボソームを持っている。細菌(原核生物)は少し小さく、30Sサブユニットと50Sサブユニットの2つから構成される70S リボソームを持っている。真核生物のミトコンドリアは細菌由来の細胞内小器官で、ミトコンドリア固有のリボソームは28Sと39Sの2つのサブユニットから構成される55S リボソームを持っている。
ヒトなどの真核生物と細菌ではリボソームの構造が異なるため、細菌のリボソームにのみ作用する特異的な阻害剤は、病原細菌に対する毒性は高くヒトに対する毒性が低い抗生物質になります。そのような抗生物質としてアミノグリコシド系化合物(ストレプトマイシン、ネオマイシン、カナマイシン)やテトラサイクリン、クロラムフェニコール、マクロライド系化合物などがあります。このような抗生物質はミトコンドリアの働きを阻害する目的で使用することもできます。
◉ ミトコンドリアのタンパク質の一部はミトコンドリアで合成される
嫌気性原始真核細胞と好気性細菌のα-プロテオバクテリアが共生を始めた初期の段階で、α-プロテオバクテリアのDNAの大半は核に移行してゲノムDNAに組み込まれました。しかし、ミトコンドリア固有の遺伝子の一部はミトコンドリア内のDNAに存在しています。
ミトコンドリアDNAは16,569bpの環状の分子で、37個の遺伝子が存在し、22個のトランスファーRNA(tRNA)と2個のリボソームRNA(rRNA)の遺伝子と、酸化的リン酸化に関与するタンパク複合体の85種類のサブユニットのうち13種類のタンパク質を作成する遺伝子が存在します。
ミトコンドリアのタンパク質の大半は核内DNA にコードされています。DNAがコードしているとは、DNAの塩基配列が特定のアミノ酸の配列を指定し、それに基づいてタンパク質が合成されることです。生物の遺伝情報はDNAに保存されており、DNAの4種類の塩基(アデニン、グアニン、シトシン、チミン)が特定の順序で並ぶことによって、タンパク質を構成するアミノ酸の順序が決定されます。この3種類の塩基の組み合わせ(トリプレット)はコドンと呼ばれ、1つのアミノ酸を指定するコードとして機能します。
核内DNA にコードされているタンパク質は、細胞質で合成された後、ミトコンドリア外膜と内膜を通過してミトコンドリア内部に輸送されて来ます。 ミトコンドリアDNA にコードされているミトコンドリア・タンパク質は、核内DNA にコードされているものと比較すると、その種類は少数です。しかし、ミトコンドリアでのタンパク質合成が阻害されると、酸素呼吸(酸化的リン酸化)ができなくなり、ミトコンドリアでのエネルギー産生が阻害されます。ミトコンドリアのリボソームのリボソームRNA(rRNA)成分は、ミトコンドリアDNAによってコードされます。一方、ミトコンドリアのリボソームタンパク質および翻訳に必要なすべてのタンパク質は、核遺伝子によってコードされ、細胞質のリボソームにより合成されてミトコンドリアに輸送されます。
ミトコンドリアのタンパク質合成は、全ての哺乳類にとって不可欠であり、酸化的リン酸化複合体の重要な成分を提供する役割を担っています。たった13種類のポリペプチドしか作られていませんが、ミトコンドリアでのタンパク合成が阻害されると細胞は生きていけません。
ミトコンドリアは核とは別に独自のDNA(mtDNA)を有しており、このmtDNAにコードされたタンパク質を合成するための独自のタンパク質合成系をもっています。
ミトコンドリアは細菌の共生が起源と考えられていて、ミトコンドリアのタンパク質合成系の仕組みは、原則的には原核細胞(細菌)のものに類似しています。つまり、哺乳類のミトコンドリア・リボソーム(55S)は、細胞質リボソーム(80S)より細菌リボソーム(70S)に近いと言えます。したがって、細菌リボソームを阻害する抗生物質は、ミトコンドリアの働きを阻害する作用があります。
◉ ドキシサイクリンとアジスロマイシンはミトコンドリアのATP産生を阻害する
ドキシサイクリンは、テトラサイクリン誘導体の抗生物質です。テトラサイクリンの有効性と安定性を改善した医薬品で、1960年代後半に米国食品医薬品局(FDA)が承認しています。グラム陽性とグラム陰性の両方の細菌に効き、にきびの治療に使われています。
テトラサイクリンは細菌のリボソームの30Sサブユニットに結合して、細菌のタンパク合成を阻害します。この細菌の30Sリボソーム・サブユニットはミトコンドリアの28Sリボソーム・サブユニットと相同性があるため、ミトコンドリアのタンパク質合成を阻害する作用もあります。この作用によってミトコンドリア機能を阻害します。
実際に、ドキシサイクリンががん細胞のミトコンドリアの働きを阻害することによって、がん細胞の増殖や腫瘍形成能を阻害する作用が多数報告されています。例えば、ドキシサイクリンはミトコンドリアの機能異常と酸化傷害を引き起こして、膠芽腫(グリオブラストーマ)の抗がん剤に対する感受性を亢進することが報告されています。 この実験では、ドキシサイクリンはミトコンドリアのATP産生を低下させ、ミトコンドリアの活性酸素種の産生量を高め、細胞内のタンパク質と脂質の過酸化を増やしました。これは、ドキシサイクリンはミトコンドリアにおけるエネルギー産生を破綻させ、酸化ストレスを高めて細胞の酸化傷害によってグリオブラストーマ細胞を死滅させることを示しています。また、ドキシサイクリンが、乳がん幹細胞の幹細胞特性の維持と上皮-間葉移行(上皮細胞が線維芽細胞のような間葉系細胞の性質を獲得して周囲組織への浸潤性が高まること)を阻止する作用が報告されています。そのメカニズムとしてミトコンドリアの働きを阻害する作用を指摘しています。(文献35)
ミトコンドリアはエネルギー(ATP)産生だけでなく、細胞内カルシウムの恒常性維持、アポトーシス制御、細胞内シグナル伝達系など多くの機能において重要な働きを担っています。ミトコンドリアが破綻して機能が阻害されると、がん細胞は生存できなくなります。
アジスロマイシンはマクロライド系の抗生物質で、細菌の50Sリボソームに結合してmRNAからタンパク質への合成を阻害します。細菌の50Sリボソームはミトコンドリアの39Sリボソームと相同性が高いので、ミトコンドリアのタンパク質合成を阻害します。
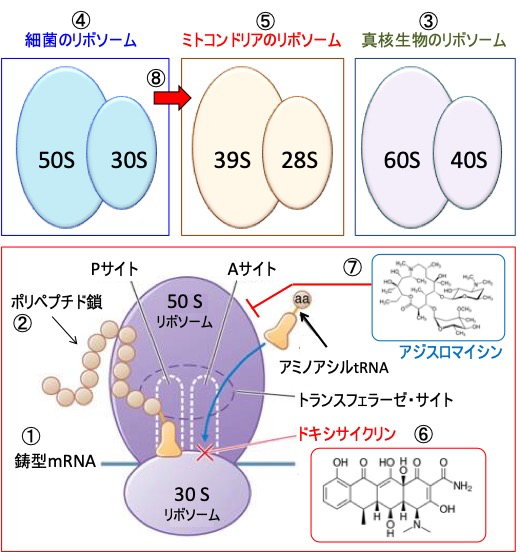
図:リボソームはmRNAの遺伝情報を読み取って(①)タンパク質(ポリペプチド鎖)へと変換する(②)。リボソームは大小2種類のサブユニットから構成され、真核生物のリボソームは40Sサブユニットと60Sサブユニットから構成される(③)。細菌は少し小さく、30Sサブユニットと50Sサブユニットの2つから構成される(④)。ミトコンドリアは細菌由来の細胞内小器官で、ミトコンドリア固有のリボソームは28Sと39Sの2つのサブユニットから構成される(⑤)。抗生物質のドキシサイクリンは細菌のリボソームの30Sサブユニットに結合して、ポリペプチド鎖へのアミノ酸の結合を阻害して、細菌のタンパク合成を阻害する機序で抗菌作用を発揮する(⑥)。アジスロマイシンは50Sリボソームに作用してタンパク質合成を阻害する(⑦)。ミトコンドリアは細菌に由来するため、細菌のリボソームの30Sサブユニットと50Sサブユニットは、それぞれミトコンドリア・リボソームの28Sサブユニットと39Sサブユニットと相同性がある(⑧)。したがって、ドキシサイクリンとアジスロマイシンはミトコンドリアのタンパク質合成を阻害する作用もあり、この作用によってミトコンドリア機能やミトコンドリア新生を阻害する。
◉ がんを感染症のように治療する
ミトコンドリアのリボソームの小サブユニットを阻害するドキシサイクリンと、大サブユニットを阻害するアジスロマイシンとの2種類の抗生物質とビタミンCを併用すると、極めて低用量でがん幹細胞を根絶できることが報告されています。
乳がん細胞株を使用した実験で、ドキシサイクリン(1μM)とアジスロマイシン(1μM)およびビタミンC(250μM)の組み合わせで治療すると、がん幹細胞の増殖が90%以上阻害されました。
ビタミンCは穏やかな酸化促進剤として働き、フリーラジカルを生成し、ミトコンドリアの新生(増生)を誘導します。この状態でミトコンドリアのリボソームのタンパク合成を阻害するとエネルギー(ATP)産生が阻害され、がん幹細胞が死滅するというメカニズムです。
ここで使用される抗生物質の濃度は、抗菌目的で使用されるドキシサイクリンおよびアジスロマイシンの濃度より低いので、抗生物質耐性菌出現に関連する潜在的な問題を回避できます。臨床試験も行われており、ドキシサイクリンが早期乳がん患者のがん幹細胞を効果的に減少させる結果が報告されています。この臨床試験では、ドキシサイクリンは、手術前の14日間、200 mgの1日量で経口投与されました。15人の患者のうち6人が対照(治療なし)で、9人がドキシサイクリンで治療されました。
ホルマリン固定パラフィン包埋サンプルの免疫組織化学分析によって、がん幹細胞、ミトコンドリア、細胞増殖、アポトーシス、および血管新生のバイオマーカーが解析されました。各患者について、術前生検標本と外科切除標本の両方で分析を実施し、ドキシサイクリン投与前から治療後までの変化が統計的に比較されました。
その結果、ドキシサイクリン投与前の生検サンプルと比較して、ドキシサイクリン投与後の腫瘍サンプルは、がん幹細胞のマーカーのCD44の統計的に有意な減少を認めました。具体的には、ドキシサイクリンで治療された患者9人中8人でCD44レベルが17.65〜66.67%低下しました。一人の患者はCD44の15%の増加を示しましたが、全体として約90%の症例においてがん幹細胞の性質の減少を認めました。
別のがん幹細胞のマーカーであるアルデヒド脱水素酵素でも同様の結果が得られました。つまり、ドキシサイクリンが生体内で乳がん患者のがん幹細胞を選択的に根絶できることを示唆しています。以上のように、細菌のリボソームを阻害して抗菌作用を発揮する抗生物質は、ミトコンドリアのリボソームの働きを阻害する副作用を持ちますが、この作用をがん治療に利用できるというアイデアです。がんを感染症にように治療するというアイデアとも言えます。
抗がん剤治療や高濃度ビタミンC点滴を行っているときに、ドキシサイクリンとアジスロマイシンの併用を試してみる価値はあります。アルテスネイトと鉄剤と5-アミノレブリン酸を組み合わせるフェロトーシス誘導療法に、さらにドキシサイクリンとアジスロマイシンを追加すると、抗腫瘍効果を増強できます。副作用が出ない低用量で効果が期待でき、しかも安価です。
ドキシサイクリンやアジスロマイシンに関するご質問やお問い合わせは、メール(info@f-gtc.or.jp)か電話(03-5550-3552)でご連絡ください。