がん細胞のフェロトーシス誘導療法のまとめ
がん細胞に選択的かつ強力にフェロトーシスを誘導するためには、複数のメカニズムをターゲットにする必要があります。がん細胞に選択的に活性酸素の産生を可能な限り高め、脂質過酸化を防いでいる抗酸化システムを十分に弱体化させ、細胞膜の過酸化脂質を増やすことが、フェロトーシス誘導に必要です。そのメカニズムを整理しています。
◉ 鉄代謝の観点からのフェロトーシス誘導療法のまとめ
がん細胞は鉄の取り込みが多く、がん細胞内にはフリーの2価鉄(Fe2+)が多く存在するというのが、がん細胞に選択的にフェロトーシスを誘導できる根拠です。したがって、がん細胞にフェロトーシスを誘導するときの中心はアルテスネイト+5-アミノレブリン酸(5-ALA)+鉄剤です。これに抗酸化力を阻害する方法と酸化ストレスを高める方法を併用すれば、フェロトーシス誘導を増強できます。
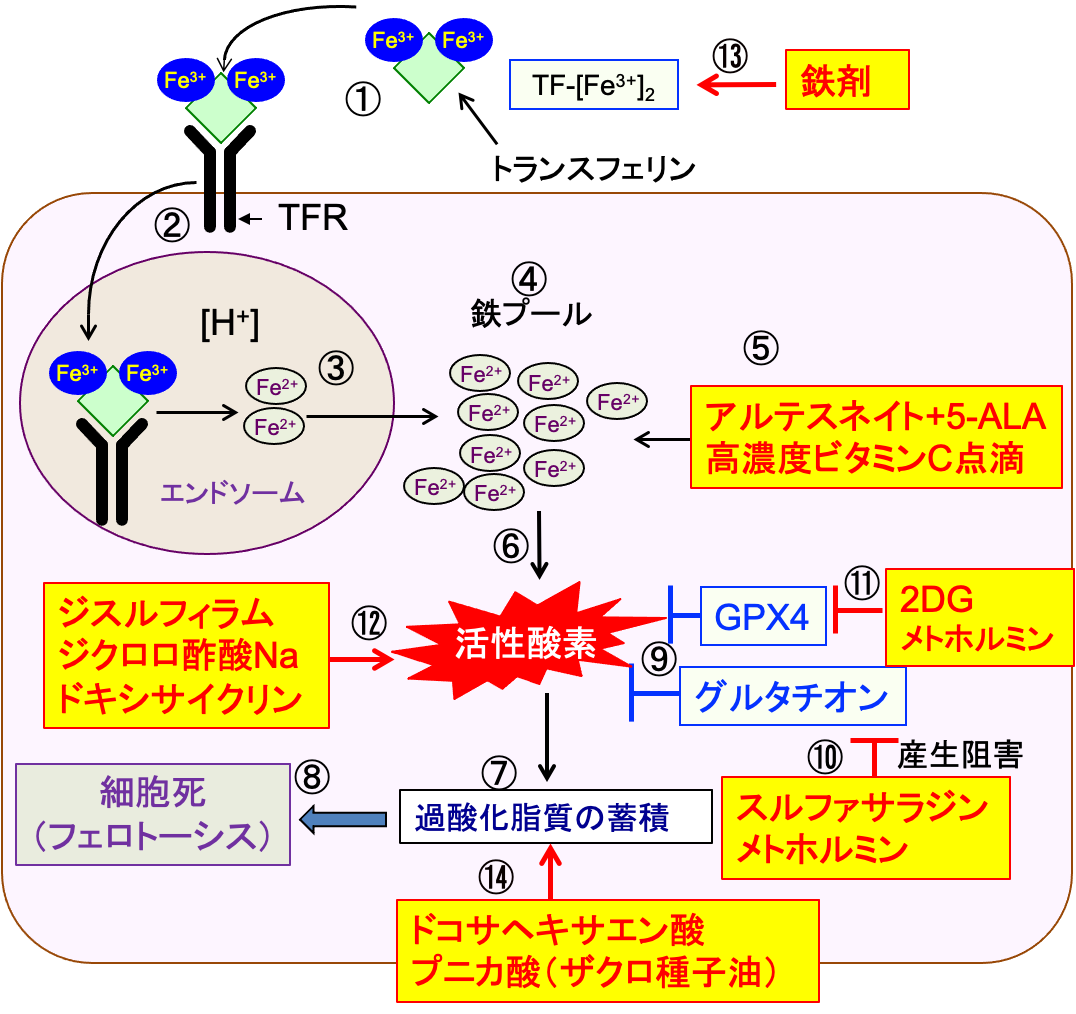
図:トランスフェリン(TF)は3価の鉄イオン(Fe3+)を運搬する(①)。細胞膜のトランスフェリン受容体(TFR)にトランスフェリンが結合すると細胞内に取り込まれる(②)。エンドソーム内の酸性の環境では鉄イオンはトランスフェリンから離れ、3価の鉄イオン(Fe3+)は2価の鉄イオン(Fe2+)に還元される(③)。2価の鉄イオンは細胞質に移行し、細胞内の様々な目的で使用される(④)。アルテスネイト+5-アミノレブリン酸(5-ALA)と高濃度ビタミンC点滴(⑤)はがん細胞内の2価の鉄イオン(Fe2+)と反応して活性酸素を発生し(⑥)、過酸化脂質の蓄積を引き起こし(⑦)、フェロトーシスによる細胞死を誘導する(⑧)。がん細胞はグルタチオンやグルタチンペルオキシダーゼ4(GPx4)の活性を高めて活性酸素を消去する(⑨)。スルファサラジンとメトホルミンはシスチン・トラスポーターの働きを阻害してグルタチオンの合成を阻害する(⑩)。2-デオキシ-D-グルコース(2-DG)とメトホルミンはATPとNADPHの産生を減らしてグルタチンペルオキシダーゼ4(GPx4)の活性を低下する(⑪)。ジスルフィラムとジクロロ酢酸ナトリウムとドキシサイクリンは活性酸素の産生を増やす(⑫)。鉄剤の投与はがん細胞内の鉄を増やしてフェロトーシスを促進する(⑬)。ドコサヘキサエン酸とプニカ酸(ザクロ種子油)は細胞膜に取り込まれ、細胞膜の脂質過酸化を促進する(⑭)。これらは、がん細胞のフェロトーシス誘導において相乗効果を発揮する。
◉ 脂質過酸化の促進の観点からのまとめ
シスチン・トランスポーター/グルタチオン/グルタチオンペルオキシダーゼ4(GPx4)による抗酸化システムの阻害は、がん細胞の細胞膜の脂質の過酸化を促進します。
2-デオキシ-D-グルコースやメトホルミンでペントースリン酸経路を阻害すると還元剤のNADPHの産生を阻害できます。
スルファサラジンはシスチン・トランスポーターを阻害します。酸化ストレスを高めるジスルフィラムの抗腫瘍効果は、スルファサラジンとの併用によって増強できます。
ドコサヘキサエン酸とプニカ酸(ザクロ種子油)は脂質過酸化を促進し、アルテスネイト、ジクロロ酢酸ナトリウム、高濃度ビタミンC点滴はがん細胞内の活性酸素とフリーラジカルの産生を増やし、酸化ストレスを高めます。
ドキシサイクリンとアジスロマイシンは、ミトコンドリアの呼吸酵素鎖のタンパク質合成を阻害してATP産生を阻害します。これらを組み合わせるとがん細胞に選択的にフェロトーシスによる細胞死を誘導できます。
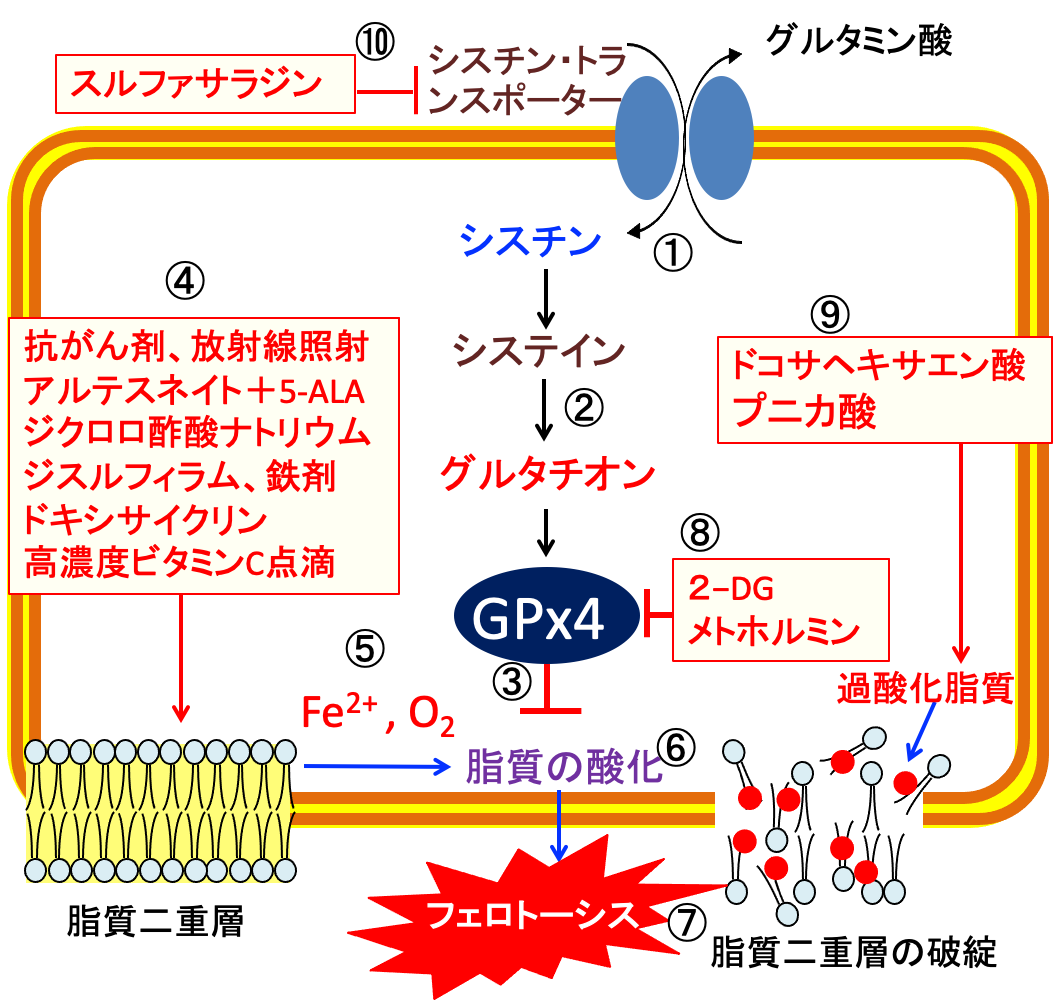
図:シスチン・トランスポーターは細胞外のシスチンを細胞内に輸送する(①)。シスチンはシステインに変換されてグルタチオンの合成が増える(②)。グルタチンペルオキシダーゼ4(GPx4)はグルタチオンを使って細胞膜の脂質の酸化を防ぐ(③)。抗がん剤、放射線照射、アルテスネイト+5-ALA(5-アミノレブリン酸)、ジクロロ酢酸ナトリウム、ジスルフィラム、鉄剤、ドキシサイクリン、高濃度ビタミンC点滴は活性酸素の産生を増やし(④)、2価の鉄イオン(Fe2+)と酸素(O2)が介在した機序で(⑤)、脂質酸化を促進し(⑥)、脂質二重層の破綻によってフェロトーシスによって死滅する(⑦)。2-デオキシ-D-グルコース(2-DG)とメトホルミンはATPとNADPHの産生を減らしてグルタチンペルオキシダーゼ4(GPx4)の活性を低下する(⑧)。細胞膜に取り込まれたドコサヘキサエン酸とプニカ酸(ザクロ種子油)は脂質過酸化を促進する(⑨)。スルファサラジンはシスチン・トランスポーターの働きを阻害してグルタチオンの合成を阻害する(⑩)。これらを組み合わせると、がん細胞に選択的にフェロトーシスによる細胞死を誘導できる。
◉ メトホルミン+2-デオキシ-D-グルコース+ジクロロ酢酸ナトリウム+ケトン食の相乗効果
2-デオキシ-D-グルコース(2-DG)は、解糖系を阻害することによって乳酸とATPの産生を阻害します。さらにペントースリン酸経路を阻害して物質合成を阻害します。
経口糖尿病薬のメトホルミンはミトコンドリアの呼吸酵素を阻害してATP産生を阻害する作用があり、さらに2-DGと同様に解糖系酵素のヘキソキナーゼの活性を阻害します。
したがって、2-DGとメトホルミンを併用すると、がん細胞のエネルギー産生と物質合成を阻害する効果を相乗的に高めることができます。エネルギー産生と物質合成の阻害は抗酸化システムを弱体化し、フェロトーシスを起こしやすくします。
メトホルミンには乳酸アシドーシスを引き起こす副作用があります。乳酸が増えて血液が酸性になる状態です。大きながん組織があると乳酸の産生が増えています。乳酸アシドーシスを防ぐために、肝臓では乳酸をグルコースに変換する糖新生が亢進します。メトホルミンは糖新生を阻害する作用があるので、乳酸産生の増加した状態でメトホルミンを服用すると、乳酸アシドーシスを起こしやすくなります。
この場合、がん細胞の解糖系を抑制し、ミトコンドリアでの酸素呼吸を増やす2-デオキシ-D-グルコースやジクロロ酢酸ナトリウムやケトン食を併用するとメトホルミンによる乳酸アシドーシスの発生を防ぐことができます。特にジクロロ酢酸ナトリウムは乳酸アシドーシスの治療に古くから使用されています。
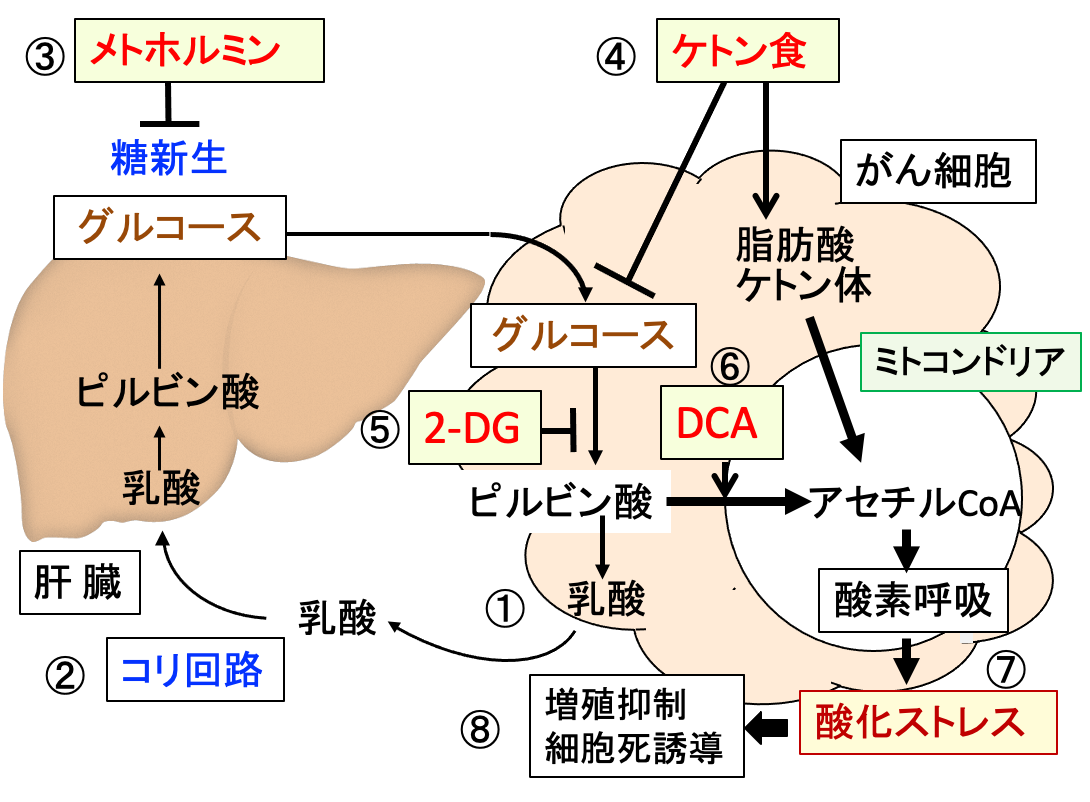
図:がん細胞は乳酸の産生が増えている(①)。乳酸によるアシドーシス(酸性血症)を防ぐため、肝臓で乳酸をグルコースに変換する。これをコリ回路という(②)。メトホルミンは糖新生を阻害するので、乳酸アシドーシスの副作用を起こしやすい(③)。ケトン食はグルコースの利用を阻害し、脂肪酸とケトン体はミトコンドリアの酸素呼吸(酸化的リン酸化)を亢進する(④)。2-デオキシ-D-グルコース(2-DG)はがん細胞の解糖系を阻害し(⑤)、ジクロロ酢酸ナトリウム(DCA)はピルビン酸脱水素酵素を活性化してピルビン酸からアセチルCoAへの変換を促進する(⑥)。その結果、2-DGとDCAはメトホルミンによる乳酸アシドーシスを防ぎ、活性酸素の産生を高めて酸化ストレスを亢進する(⑦)。これらは相乗効果で、がん細胞の増殖を抑制できる(⑧)
ケトン食だけでは抗腫瘍効果は弱いのですが、2-デオキシ-D-グルコースとメトホルミンとジクロロ酢酸ナトリウムを併用すると、がん細胞の増殖を抑制できます。さらに抗酸化システムを阻害するジスルフィラムやスルファサラジンを併用すると、がん細胞を酸化ストレスで自滅できます。
2-デオキシ-D-グルコースとジクロロ酢酸ナトリウム(+ビタミンB1+R体αリポ酸)とメトホルミンを併用したがん治療の有効性のメカニズムを下の図にまとめています。
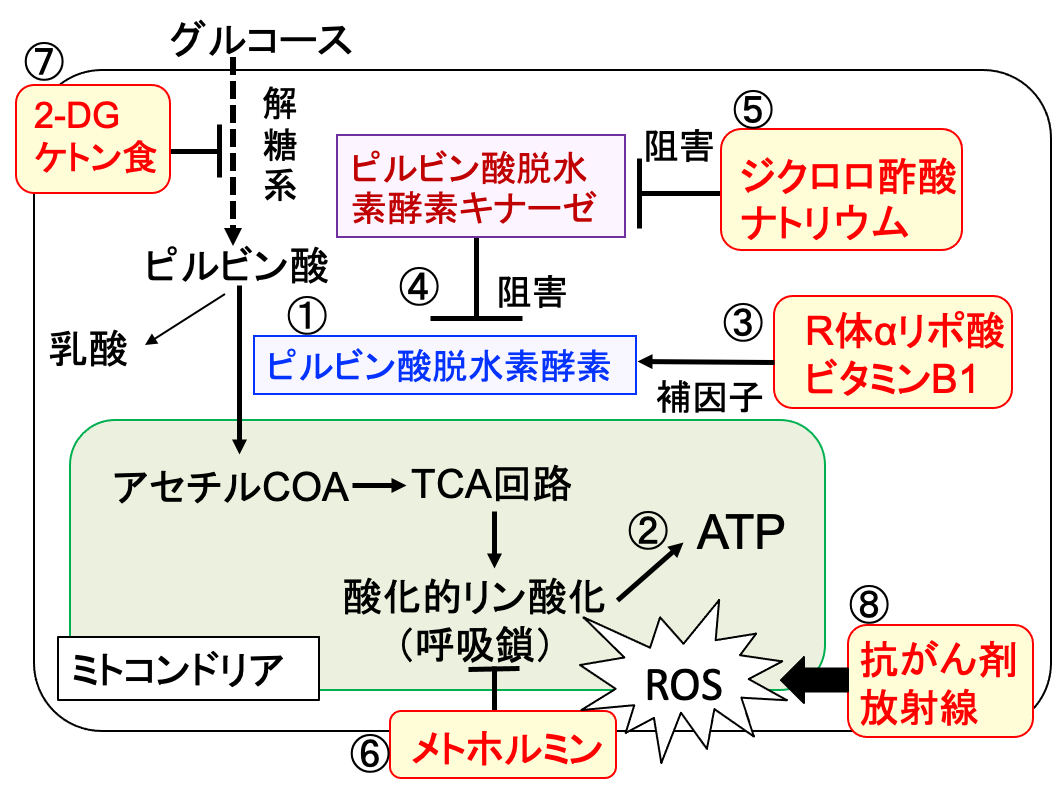
図:グルコースが解糖系でピルビン酸に変換された後、ピルビン酸脱水素酵素(①)によってアセチルCoAに変換される。アセチルCoAはミトコンドリア内でTCA回路と呼吸酵素複合体における酸化的リン酸化によってATPが産生される(②)。R体αリポ酸とビタミンB1はピルビン酸脱水素酵素の補因子として働く(③)。ピルビン酸脱水素酵素はピルビン酸脱水素酵素キナーゼによってリン酸化されることによって活性が阻害される(④)。ジクロロ酢酸ナトリウムはピルビン酸脱水素酵素キナーゼを阻害してピルビン酸脱水素酵素を活性化する(⑤)。メトホルミンは呼吸酵素複合体Iを阻害してミトコンドリアでの活性酸素の産生を増やす(⑥)。2-デオキシ-D-グルコース(2-DG)とケトン食は解糖系を阻害する(⑦)。抗がん剤や放射線治療は活性酸素の産生を増やす(⑧)
抗がん剤治療や放射線治療を行うときに、2-デオキシ-D-グルコース、ジクロロ酢酸ナトリウム、R体αリポ酸、ビタミンB1、メトホルミン、ケトン食を併用すると、抗腫瘍効果を増強できます。
アルテスネイトを用いたフェロトーシス誘導の促進にも有効です。この組み合わせはがんの補完・代替療法として今までに多くの患者さんに行っていますが、副作用はほとんど経験せず、顕著な有効性を確認しています。
「がんのフェロトーシス誘導療法」に関するご質問やお問い合わせは、メール(info@f-gtc.or.jp)か電話(03-5550-3552)でご連絡ください。